Cerchiamo di comprendere quali sono i meccanismi che gestiscono il movimento lungo l’Apparato Digerente.
L’attivita’ motoria dei vari organi del sistema gastrointestinale e’ organizzata in modo da garantire la specifica funzione di ciascun singolo organo. Questo comporta che l’attivita’ motoria dell’esofago sia diversa da quella dello stomaco, che a sua volta differisce da quella dell’intestino. Inoltre è organizzata in modo da ottimizzarne il comportamento per le differenti condizioni fisiologiche (periodo prandiale e digiuno) e per azioni particolari come il vomito (meccanismo della retropulsione).
Ricordiamo che la muscolatura e’ striata nella bocca, nella faringe, nell’esofago superiore e nel pavimento pelvico (sfintere anale esterno), mentre a tutti gli altri livelli del sistema gastrointestinale e’ presente muscolatura liscia di tipo viscerale.
A quest’ultima sono associate cellule specializzate (non nervose) a funzione pacemaker dette cellule interstiziali di Cajal localizzate all’interfaccia fra gli strati longitudinale e circolare della tonaca muscolare e sulla superficie submucosale dello strato muscolare circolare (dette regioni pacemaker).
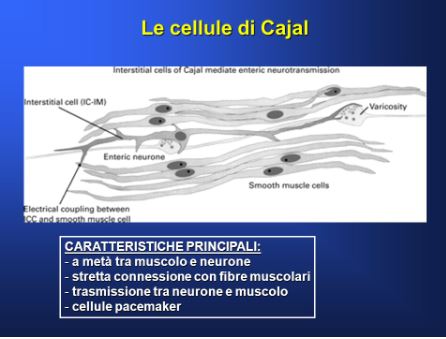
Hanno un potenziale di riposo di membrana intorno a -50 mV ( differenza di potenziale elettrico, misurabile in una cellula, tra l’interno che presenta cariche prevalentemente negative e lo spazio extracellulare che presenta cariche prevalentemente positive) non stabile con fluttuazioni ritmiche dovute alle oscillazioni della concentrazione di calcio all’interno della cellula. Questa instabilità genera ritmicamente le cosiddette onde elettriche lente o ritmo elettrico basale (REB) ovvero onde di depolarizzazione (diminuzione del valore assoluto del potenziale di riposo di membrana di una cellula che genera un potenziale elettrico) che sono alla base dell’attivita’ contrattile spontanea della muscolatura liscia viscerale. Le cellule interstiziali di Cajal sono interconnesse fra loro e con le fibre muscolari lisce della parete viscerale mediante gap junction, formando un sincizio elettrico funzionale. Le onde elettriche lente generate si propagano passivamente, attraverso le gap junction, alle fibre muscolari lisce degli strati di muscolatura sia circolare sia longitudinale diffondendosi, quindi, in tutto il tessuto muscolare di un distretto. Tuttavia, pur essendo onde di eccitabilità, non determinano contrazione efficace perché non portano mai il potenziale di membrana di riposo delle cellule muscolari lisce al potenziale soglia, per cui avviene solo una depolarizzazione che a sua volta produce solo delle piccole contrazioni dette toniche.(ONDE LENTE 3-18/min).
La frequenza delle onde elettriche lente capaci di produrre contrazioni efficaci varia lungo il tubo gastroenterico: Nello stomaco distale (antro gastrico) la frequenza delle onde lente non supera le tre contrazioni al minuto. Nell’intestino tenue, la frequenza delle onde elettriche lente diminuisce progressivamente andando dal duodeno (dove la frequenza e’ massima e ha il valore di 16-18 onde/min) all’ileo terminale (dove la frequenza e’ minima e ha il valore di 8 onde/min). Il gradiente che si viene a creare favorisce pertanto lo spostamento in direzione aborale del contenuto luminale. Nell’intestino crasso, la frequenza delle onde elettriche lente e’ minima nel cieco e colon prossimale (6 onde/min) ed e’ invece massima nel colon sigmoideo (10-16 onde/min ). Questo gradiente invertito della frequenza delle onde elettriche lente rispetto a quello esistente nell’intestino tenue favorisce il prolungato rimescolamento del contenuto luminale a livello dell’intestino crasso ottimizzando la disidratazione e il compattamento delle feci.
Solo se un’onda elettrica lenta supera il valore soglia(-40mV) si possono generare nelle cellule muscolari lisce uno o piu’ potenziali d’azione, cui si associa contrazione delle fibre muscolari. In assenza di potenziali d’azione non si hanno contrazioni. In sostanza possiamo dire che esiste un ritmo elettrico basale sempre attivo provocato dalla depolarizzazione delle cellule muscolari lisce indotte dalle frequenza delle onde elettriche lente ma che per ottenere delle contrazioni valide è necessario che si verifichino dei potenziali d’azione che però non sempre si associano a tutte le onde lente. Infatti le contrazioni provocate dai potenziali d’azione avvengono solo quando i neuroni inibitori del SNE di quel segmento non sono attivi; in altri termini i motoneuroni inibitori controllano il ritmo elettrico basale impedendo la creazione di potenziali d’azione e quindi le contrazioni, ma una volta bloccati sono in grado di dare inizio a una contrazione determinando anche la distanza e la direzione di propagazione della stessa una volta iniziata. Questi motoneuroni inibitori della muscolatura circolare sono in stato di continua attività, per cui la contrazione muscolare può avere luogo solo quando essi vengono inibiti a opera di interneuroni dei circuiti di controllo del SNE. Solo a livello degli sfinteri, i motoneuroni inibitori sono invece normalmente inattivi.
La creazione dei potenziali d’azione porta alla formazione di una attivita’ motoria nel digerente che si manifesta sotto tre forme:
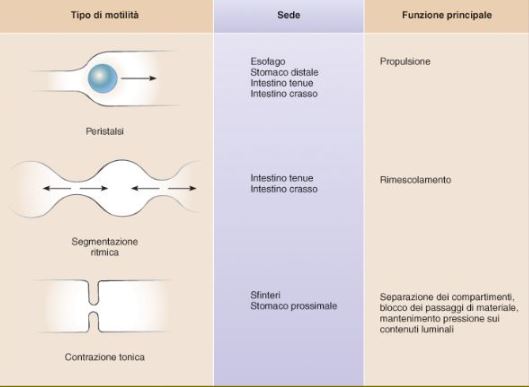
La PERISTALSI: e’ un’onda di contrazione il cui risultato finale e’ la propulsione in direzione aborale del contenuto del lume. Durante l’attività peristaltica, la muscolatura della parete gastrointestinale presenta un comportamento stereotipato determinato dall’attività del SNE. Nella fattispecie, nel segmento a valle del contenuto luminale che sta progredendo, lo strato di fibre muscolari longitudinali si contrae, mentre lo strato di fibre muscolari circolari si rilascia: il tutto provoca un accorciamento dell’asse longitudinale del segmento cui si associa l’aumento del suo diametro con espansione del lume. Questa è l’attività del segmento recettivo, cioe’ di quello che si prepara ad accogliere il materiale luminale in progressione. Contemporaneamente, nel segmento a monte del contenuto luminale in progressione, si verifica la contrazione dello strato di fibre muscolari circolari, mentre lo strato di fibre muscolari longitudinali si rilascia. E’ questa l’attività del segmento propulsivo, cioè di quello che spinge il contenuto luminale verso il segmento recettivo. I segmenti inizialmente a valle del fronte di avanzamento diventano quindi segmenti recettivi, per diventare poi a propria volta segmenti propulsivi, garantendo in questo modo il progressivo spostamento dell’attività peristaltica (organizzata appunto in segmenti propulsivi e recettivi) lungo il tubo gastroenterico. La formazione dei segmenti propulsivo e recettivo (cioe’ il riflesso peristaltico) è gestita a livello locale dal SNE che crea dei veri archi riflessi neurologici intrinseci locali. Il circuito di base di questi archi e’ costituito dalle connessioni sinaptiche tra neuroni sensoriali intrinseci, interneuroni e neuroni motori che a loro volta possono subire dei blocchi da parte di neuromotori inibitori. La distanza percorsa dalla propulsione peristaltica e’ quindi determinata dal numero di blocchi di questi circuiti lungo l’intestino.
LA SEGMENTAZIONE RITMICA: sono contrazioni ritmiche dello strato muscolare circolare, finalizzate a rimescolare e dividere il contenuto luminale;
LA CONTRAZIONE TONICA:sono contrazioni prolungate, presenti essenzialmente nella porzione prossimale dello stomaco e negli sfinteri. Possono durare per minuti od ore.
Le tre componenti dell’attività motoria sono anche gestite da archi riflessi neurologici estrinseci di tipo breve, in cui il SNE invia impulsi al sistema nervoso autonomo(in linea di massima, il sistema parasimpatico aumenta l’ampiezza delle onde elettriche lente, mentre quello simpatico la diminuisce) e di tipo lunghi in cui gli impulsi prevalentemente sensitivi vengono inviati al SNC.
Ultima componente che interviene nella regolazione della motilità sono le strutture endocrine intrinseche del tubo digerente
Vediamo la motilità delle singole componenti del tubo digerente:
Bocca:
E’ costituita da una sequenza di fasi (volontarie e riflesse) coordinate dal centro della deglutizione bulbare cerebrale. La fase orale è costituita da:
1) Masticazione: gli alimenti sono finemente triturati, aumentandone così la superficie di attacco, su cui possono agire gli enzimi digestivi. In particolare, a livello orale l’amido cotto è digerito dall’amilasi salivare. Inoltre, la liberazione di aromi dagli alimenti ingeriti stimola i chemiocettori orali e olfattivi, favorendo le secrezioni salivare, gastrica e pancreatica. La stimolazione tattile all’interno della bocca determina a seconda della quantità degli alimenti presenti un aumento o una diminuzione omolaterale della contrazione dei muscoli che tengono chiusa la mandibola favorendo il passaggio da una guancia all’altra del materiale alimentare. Il centro bulbare cerebrale coordina questi movimenti e quelli della lingua spingendo il tutto verso i denti. È importante ricordare che i denti esercitano una forza notevole (circa10 kg) sulle sostanze presenti nella bocca.
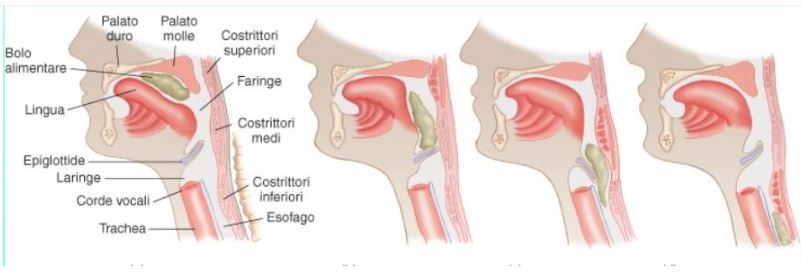
2) Deglutizione: La fase propulsiva avviene durante la masticazione quando il bolo è spinto verso la faringe ad opera della lingua. La punta della lingua separa un bolo di 5-15 ml dal resto del materiale contenuto nella bocca e lo porta all’indietro. Il palato molle si solleva e chiude il passaggio con le vie nasali; la mandibola è chiusa; la contrazione della lingua proietta il bolo contro la parete faringea posteriore; la laringe si innalza e l’ epiglottide si abbassa, evitando il rigurgito del bolo verso la vie aeree ed impedendo quindi il soffocamento; infine, lo sfintere faringo-esofageo si rilascia per permettere il passaggio del bolo nell’esofago. Quando il bolo alimentare ha raggiunto lo sfintere superiore dell’esofago, l’epiglottide si innalza e la laringe si abbassa. In tal modo si ristabilisce la comunicazione con le vie respiratorie e la respirazione può avere nuovamente luogo.
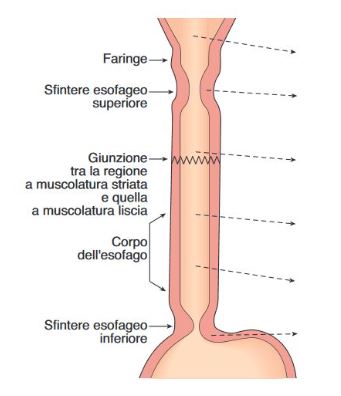
Esofago: L’esofago assicura il trasporto del bolo alimentare allo stomaco. All’estremità superiore si trova lo sfintere faringo-esofageo(o sfintere superioredell’esofago) e alla sua estremità inferiore lo sfintere gastro-esofageo o cardias, che supera il diaframma attraverso lo iato. Tra i due è compreso il corpo dell’esofago che si estende per circa 20 centimetri. La muscolatura del quarto superiore dell’esofago è striata, con innervazione somatica, mentre quella del terzo inferiore è liscia, con innervazione autonoma. L’avanzamento del bolo è dovuto all’azione peristaltica coordinata dei diversi segmenti dell’esofago. Lo sfintere faringo-esofageo a riposo è contratto tonicamente e possiede una pressione endoluminale di circa 60 mmHg. Questa pressione limita la penetrazione di aria nell’esofago nel corso della respirazione e previene il rigurgito esofago-faringeo. Al momento della deglutizione la muscolatura dello sfintere faringo-esofageo si rilascia. Il bolo penetra nell’esofago, favorito dal fatto che anche il corpo dell’esofago è rilasciato, Subito dopo lo sfintere si contrae nuovamente. Il bolo, penetrato nell’esofago, è spinto da un’onda peristaltica, che viaggia alla velocità di 2-4 cm/sec e determina un’onda pressoria di 30-120mmHg. La peristalsi del corpo è coordinata dall’innervazione intrinseca. Dopo il passaggio dell’onda peristaltica, per riflesso neurologico locale, possono insorgere ulteriori onde peristaltiche (peristalsi secondaria), la cui funzione è di permettere al materiale ancora presente nell’esofago di raggiungere lo stomaco(azione di pulizia). Lo sfintere gastro-esofageo o cardias è costituito da un ispessimento della muscolatura circolare a livello della giunzione gastro-esofagea, di circa 3-6 cm . Quando è a riposo lo sfintere esercita una contrazione tonica con una pressione endoluminale di circa 40mmHg. Quando il bolo si avvicina allo sfintere, esso si rilascia consentendo il passaggio del bolo nello stomaco.
Stomaco
Presenta due tipi di motilità:
Durante il digiuno: è una motilità interprandiale gestita dai complessi motori migranti(MMC). In pratica si verificano dei periodi di quiescenza alternati a contrazioni piuttosto intense (crampi da fame), generate dal pace-maker gastrico, che si propagano dallo stomaco verso l’intestino. Hanno una funzione di pulizia in quanto svuotano lo stomaco dai residui del pasto precedente. Sono presenti sia durante la veglia che durante il sonno e cessano all’arrivo del bolo alimentare.
Durante il pasto: al suo arrivo possiamo identificare nello stomaco tre porzioni con competenza funzionale diversa:
Il fondo:non ha un ruolo motorio significativo ma, grazie al rilasciamento della propria muscolatura, aumenta il volume gastrico, per evitare che la pressione intragastrica aumenti in maniera significativa all’arrivo del bolo alimentare il che potrebbe essere dannoso. Non ospita cibo ma gas.
Il corpo: esegue un rimescolamento, cioè fa diventare il bolo chimo gastrico (poltiglia mescolata con i succhi gastrici; prima del corpo è ancora bolo alimentare). Così avviene il primo atto digestivo, è la zona con maggiore volume. Le sue contrazioni dette di segmentazione non sono propulsive ma hanno lo scopo di segmentare e di rimescolare il cibo contenuto nello stomaco, in modo che possa essere efficacemente mescolato agli enzimi digestivi. Trattasi di contrazioni anulari che, a differenza di quelle peristaltiche, non si propagano. La durata di questa attività motoria varia a secondo della quantità di cibo ingerito e dal suo stato chimico-fisico . L’acqua ha uno svuotamento rapidissimo ed è tanto più veloce quanto maggiore è il suo volume. Al contrario in un pasto solido, la velocità di rimescolamento dipende dalla grandezza delle particelle. Se un cibo è prevalentemente glucidico si ha rapido rimescolamento e uno svuotamento quasi totale nell’arco della prima ora (in 30-40 min la maggior parte del glucosio è stato già mobilizzato) , se è proteico è più lungo, così come un pasto grasso. E’ un attività che non eleva di molto la pressione e non chiude mai il lume gastrico.
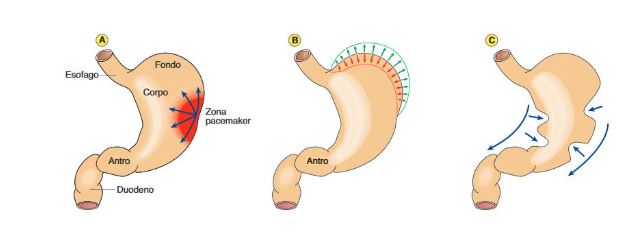
A riempimento raggiunto e soprattutto una volta formatosi il chimo iniziano onde di contrazione peristaltiche a partenza da una zona pacemaker, localizzata nel punto di passaggio tra fondo e corpo lungo la grande curvatura. Qui si ha depolarizzazione spontanea che poi si propaga all’antro.
L’antro, che ha volume minore, con una prevalenza della muscolatura circolare, il cui spessore è maggiore rispetto a quello degli altri segmenti. Presenta un REB max di 3 onde al minuto che come abbiamo visto inizia nella zona pace-maker. Inizialmente le onde di contrazione appaiono deboli ma aumentano di forza man mano che si avvicinano alla valvola pilorica che è chiusa. La frequenza del REB è uguale in tutto lo stomaco (3cicli/min), ma la sua velocità aumenta dal corpo (0,5cm/sec) all’antro (4 cm/s). Ciò significa che, considerando la lunghezza dell’antro (circa 4 cm), in un secondo il REB ne determina la contrazione, Questa regione serve a incanalare il chimo gastrico verso lo sfintere pilorico e seleziona il contenuto gastrico in termini di frazionamento( solo i frammenti < 2 mm vengono spinti attraverso il piloro nel duodeno)e in termini qualitativi. Questa azione di incanalamento, spremitura e forzatura dello sfintere pilorico viene detta propulsione per favorire lo svuotamento gastrico (a patto che il chimo abbia pH, volume e consistenza compatibili con il duodeno).Tutti i frammenti > a 2 mm tornano invece indietro nel corpo per essere ulteriormente sminuzzati. Questo processo detto di retropulsione avviene più volte fino a quando il contenuto gastrico non viene svuotato completamente
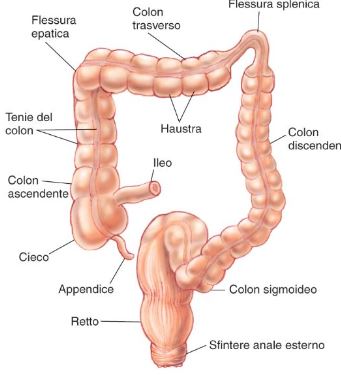
Colon:
Comunemente il materiale si ferma per 16-18 ore ed è formato da residuati di diversi pasti.
Presenta quattro tipi di movimento:
1) Austrazioni: Segmenti adiacenti di colon sono separati tra loro da contrazioni prolungate anulari della muscolatura liscia (segmentazione), poco organizzate nello spazio e nel tempo, che non si propagano o si propagano per pochi centimetri in entrambe le direzioni. Tali contrazioni determinano un incremento della pressione intraluminale che spinge il materiale presente in entrambe le direzioni (movimento pendolare). L’effetto delle contrazioni australi è quello di rimescolare il contenuto luminale, permettendo un più efficace assorbimento di acqua e sali minerali e quindi un compattamento del materiale fecale, infatti, ad ogni contrazione, la parte più distale del contenuto luminale si dirige aboralmente.
2) Movimenti peristaltici propulsivi: dopo l’assunzione di un pasto si manifestano per circa due ore in particolare nei tratti distali. Il colon presenta movimenti complessivamente volti a rimescolare il contenuto luminale e a permetterne la progressione in senso caudale. I primi predominano nelle porzioni prossimali (cieco e colon ascendente) con fenomeni di retropulsione , i secondi nelle parti distali (colon trasverso,discendente e sigmoide) con propulsione distale .
3) Movimenti di massa: nel colon trasverso e nel colon discendente si osservano i cosiddetti movimenti di massa, determinati da contrazioni della muscolatura che iniziano nella flessura epatica, si propagano ininterrottamente in senso distale e spingono il contenuto luminale verso il retto. I movimenti australi cessano prima dell’insorgenza del movimento di massa, ma riprendono quando quest’ultimo termina. I movimenti di massa precedono abitualmente la defecazione e avvengono 2-3 volte al giorno; solitamente non si presentano durante il sonno. La loro insorgenza è favorita dall’ingestione di cibo e dall’attività fisica. Durante le patologie accompagnate da diarrea sono più frequenti, mentre sono meno frequenti nei soggetti affetti da stipsi.
4) Defecazione: Come per il resto del tubo digestivo, la parete rettale include la muscolatura liscia che si suddivide in uno strato circolare interno ed uno strato longitudinale esterno. A livello dell’ano, lo strato circolare interno si ispessisce a ormare lo sfintere anale interno; Esiste anche una muscolatura striata che costituisce lo sfintere anale esterno. Quest’ultimo è il più superficiale dei muscoli del perineo ed è composto da fibre muscolari toniche, a contrazione lenta, e da fibre fasiche, a contrazione rapida. Vediamo le due fasi dell’attività motoria del tratto rettale:
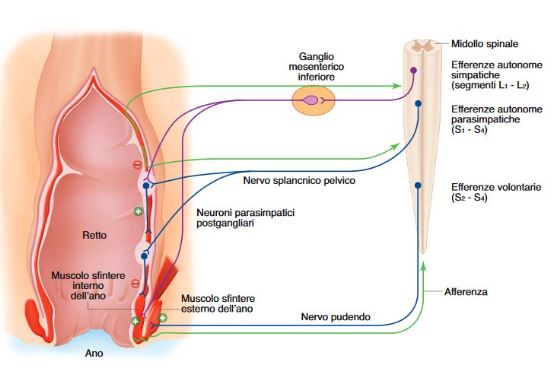
A) Continenza: La continenza è resa possibile dal colon e dal retto che agiscono da serbatoio e supporto delle feci, dagli sfinteri che ne impediscono la fuoruscita e da un sistema recettoriale che consente la percezione della presenza di materiale e permette di distinguere la presenza di gas, feci solide o liquide. A riposo lo sfintere anale interno è massimamente contratto (40-70 mmHg) per azione riflessa del simpatico (centri L1-L2 e ganglio mesenterico inferiore), ma può essere inibito per via riflessa sia dai plessi intrinseci, sia dalle efferenze autonome parasimpatiche dei centri S2-S4 e del nervo splancnico pelvico. Lo sfintere anale esterno è anch’esso tonicamente contratto (ma non massimamente) attraverso un riflesso spinale controllato dalle vie efferenti volontarie dei centri S2-S3 e dai nervi pudendi (motoneuroni somatici).
B) Defecazione: Inizia nel retto dove le pareti vengono distese dall’arrivo e ristagno delle feci, attivando dei recettori meccanici. Ciò determina rilasciamento dello sfintere interno (mediato dai plessi intramurali) e la contrazione dello sfintere anale esterno. Se l’evacuazione non avviene (per inibizione dei centri encefalici perchè impossibilitati ad andare in bagno) si ha rilasciamento recettivo del retto, con conseguente riduzione della tensione e rimozione dello stimolo, fino a quando non arrivano altre feci. L’arrivo nel retto di altro materiale fecale, facendo aumentare ulteriormente la tensione delle pareti, determina anche il rilasciamento dello sfintere esterno. perchè i motonueuroni del nervo pudendo che innervano lo sfintere esterno sono inibiti dall’attivazione dei nervi parasimpatici pregangliari. I centri corticali sono anche responsabili, durante la defecazione, del coordinamento dell’attività dei muscoli striati coinvolti: contrazione dei muscoli addominali, dei muscoli inspiratori, della glottide (manovra di Valsalva) e dei muscoli del pavimento pelvico. Tale componente potenzia la spinta verso l’esterno della massa fecale.
Dal punto di vista neurologico,come già scritto, il gradiente delle onde lente nel colon è invertito( da 6 onde/min. nel cieco a 10-16 onde/min. nel sigma). Il parasimpatico ha un effetto eccitatorio e la sua massima espressione è il riflesso gastro-colico: quando il cibo arriva nello stomaco, si osserva un aumento della motilità del crasso tale da poter provocare il riflesso della defecazione. Al contrario il simpatico ha un effetto inibitorio e il riflesso colo-colico ne è un esempio: la distensione di un tratto di intestino crasso determina una inibizione riflessa della motilità.
Per quanto riguarda il controllo ormonale, hanno attività eccitatoria sulla motilità: in particolare la gastrina, la CCK, l’insulina e la motilina; hanno invece azione inibitoria la secretina, il glucagone. Solo la gastrina e la CCK hanno effetto a dosi fisiologiche, mentre le altre sostanze hanno effetto solo a dosi superiori a quelle fisiologiche.